- What combinations of life-history traits do organisms have, and why?
- How does conflict between siblings evolve and what effects does it have on parents?
- Where should offspring be placed
- How many offspring should there be, and what sex?
- How does speciation occur and what factors affect species richness?
- Why are some organisms good dispersers and others poor dispersers?
- How often, when, and why have evolutionary changes taken place?
These seemingly disparate questions can all be answered with a knowledge of evolutionary processes in combination with a knowledge of the environment in which the organisms live, and form the field of Evolutionary Ecology. Our tools are observations, experiments and phylogenies, which provoke us to build theories, which are then tested with observations, experiments and phylogenies......
Top of the page
- What combinations of life-history traits do organisms have, and why? In many organisms, such as mammals, birds and many plants, organisms develop under physiological growth "laws" and decide when to mature, leading to a fast/slow continuum of life-history traits with many traits related strongly to body size. However many organisms, such as insects, grow to maturity on discrete packages of resource found by their parents. We have described the relationships among life-history traits in insect parasitoids, where the mode of development is a major axis of life-history variation, and very little variation is correlated with body size across species. These relationships seem to reflect the constraints imposed by finding and exploiting hosts in the environment. Body size in one family seems to evolve in response to changes in host size, but only at slow rates.
see:
Mayhew P.J. & Blackburn T.M. 1999. Does development mode organize life-history traits in the parasitoid Hymenoptera? J. Anim. Ecol. 68: 906-916.
Mayhew P.J. & Hardy I.C.W. 1998: Nonsiblicidal behavior and the evolution of clutch size in bethylid wasps. Am. Nat. 151: 419-424.
Top of the page
- How does conflict between siblings evolve and what effects does it have on parents? Sibling rivalry is common throughout the animal and plant kingdoms. Once it has evolved, tolerance may have difficulty spreading again. We have shown that tolerance can be remarkably stable in some parasitoid taxa allowing them to develop a wide range of clutch sizes, including small gregarious broods (as in bethylid wasps, shown below).

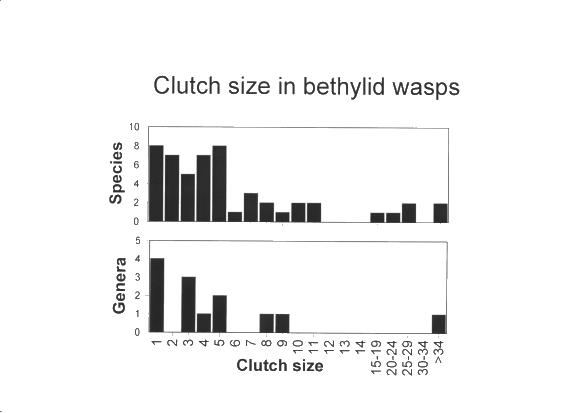
Furthermore, tolerance has repeatedly invaded intolerance, leading us to ask how this can have happened. We are currently investigating this question in the braconid genus Aphaereta, where some species have fighting larvae and develop solitarily even when several females lay eggs in a host, whilst very close relatives have tolerant larvae (see below). Recent work by John Pexton suggests that sibling rivalry has had numerous other life history consequences in Aphaereta, some of which may make affect the evolution of tolerance and intolerance.


see:
Pexton J.J. & Mayhew P.J. 2001. Immobility: the key to family harmony? Trends. Ecol. Evol. 16: 7-9.
Mayhew P.J. & van Alphen J.J.M. 1999: Gregarious development in alysiine parasitoids evolved through a reduction in larval aggression. Anim. Behav. 58: 131-141.
Mayhew P.J., Ode P.J., Hardy I.C.W. & Rosenheim J.A. 1998: Parasitoid clutch size and irreversible evolution. Ecol. Lett. 1: 139-141.
Mayhew P.J.1998: The evolution of gregariousness in parasitoid wasps. Proc. Roy. Soc. Lond. B 265: 383-389.
Mayhew P.J. & Hardy I.C.W. 1998: Nonsiblicidal behavior and the evolution of clutch size in bethylid wasps. Am. Nat. 151: 419-424.
Top of the page
-
Where should offspring be placed?
Reproducing organisms face many decisions about their offspring which should be optimized by natural selection. One of these is where to put them. We have been investigating this problem in herbivorous insects, which seem to make very poor decisions about where to place their offspring. Recent work has suggested several reasons for this, including rather uninteresting
methodological artifacts, as well as biologically interesting ones (parents may lay their eggs where it is best for themselves, rather than their offspring, to feed).
 see:
see:
Mayhew P.J. 2001. Herbivore host choice and optimal bad motherhood. Trends Ecol. Evol. 16: 165-167.
Mayhew P.J. 1998. Testing the preference-performance hypothesis in phytophagous insects: lessons from Chrysanthemum Leafminer (Diptera: Agromyzidae). Environ. Entomol. 27, 45-52
Mayhew P.J. (1997) Adaptive patterns of host-plant selection by phytophagous insects. Oikos 79, 417-428
Top of the page
-
How many offspring should there be? This is a particular problem when broods are small, when the decision is often to have one large or two small offspring. Optimal solutions to this problem should show a decrease in the variance in offspring size with increasing clutch size. This is found in the parasitic wasp,
Laelius pedatus (below).
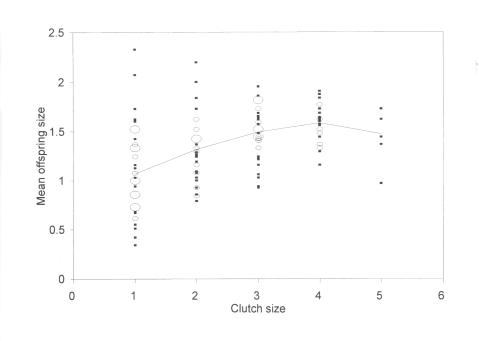
The decrease in variance is not as rapid as predicted and the mean offspring size actually increases with clutch size too (see above), a trend found in other species. We have developed models which show that strong constraints are required, such as limits on egg production rates after host encounter, to make this an optimal solution.
see:
Mayhew P.J. & Glaizot O. 2001. Integrating theory of clutch size and body size evolution for parasitoids. Oikos 92: 372-376.
Mayhew, P. J. 1998. Offspring size-number strategy in the bethylid parasitoid Laelius pedatus. - Behav. Ecol. 9: 54-59.
Top of the page
-
What sex of offspring? Some organisms, such as wasps, are able to adjust the sex ratio of their offspring in response to local conditions. In Laelius pedatus, complex sex allocation strategies occur. On very small hosts, single male eggs are laid. On slightly larger hosts, single female eggs are laid. This strategy is known as conditional sex expression. On still larger hosts, multiple-egg broods are laid which nearly always consist of one male and one or more females. This trend is also found across species within the family, and is probably due to competition between males for mates. The variation around the cross-species trend is explained by variation in the relative size of males, which may indicate the dispersal ability of the male.
see:
Mayhew P.J. & Pen. I. 2001. Comparative analysis of sex ratios Chapter 6 In Hardy I.C.W. (ed.) Sex Ratio Handbook. Cambridge University Press, in press.
Hardy I.C.W. & Mayhew P.J. 1998. Partial local mating and the sex ratio: indirect comparative evidence. Trends. Ecol. Evol. 13: 431-432.
Hardy I.C.W. & Mayhew P.J. 1998. Sex ratio, sexual dimorphism and mating structure in bethylid wasps. Behav. Ecol. Sociobiol. 42: 383-395.
Mayhew P.J. & Godfray H.C.J. 1997. Mixed sex allocation strategies in a parasitoid wasp. Oecologia 110: 218-221.
Top of the page
-
How does speciation occur and what determines species richness? Some branches of the evolutionary tree have more species than others. Why is
this? We investigated this question with Ole Seehausen (now at Hull) in African cichlid fish, which have radiated extremely rapidly and are the most species rich vertebrate family. We tested the idea that disruptive sexual selection on male nuptial coloration is responsible for the rapid diversification of these fish. The presence of nuptial coloration is associated with polygamous mating, indicative of the strength of sexual selection, and is only found in the most species rich clades. Furthermore, sister species often differ in nuptial colour, but not melanic markings (see below). Melanic markings are associated with ecological traits. The trends are consistent with the idea that disruptive mate choice drives rapid speciation in the group.
see:
Seehausen O, Mayhew P.J. & van Alphen JJM 1999. Evolution of colour
patterns in East African cichlid fish. J. evol. Biol. 12: 514-534.
Top of the page


-
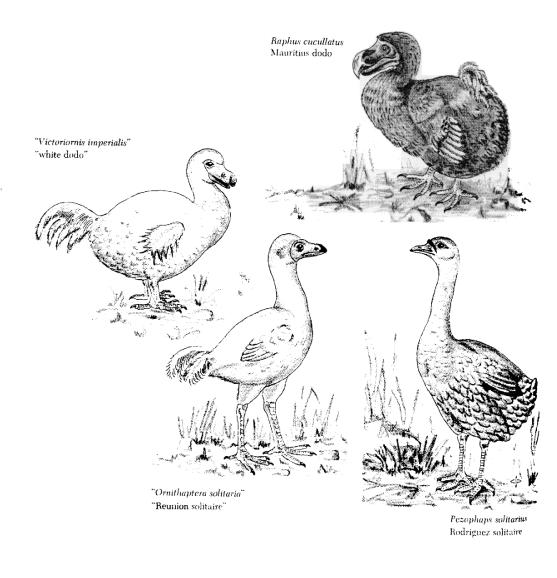
Why are some organisms good dispersers and others poor dispersers? One of the most striking correlates of dispersal ability is island occupation,
with island floras and faunas often evolving reduced dispersal, a trend famously illustrated by the flightless island birds such as the Dodo (below). Together with Calvin Dytham, we have been asking asking what promotes reduced dispersal ability on islands. Simulation models suggest that reduction in dispersal ability can rapidly evolve by loss of good dispersers at sea soon after island colonization.
see:
Dytham C & Mayhew P.J. in prep. Evolution of dispersal on islands.
Top of the page
-
How often do evolutionary changes take place? This question can be answered with a knowledge of the evolutionary relationships between groups, a knowledge
of their traits, and a model of evolution. We have been reconstructing the evolutionary relationships of bethylid wasps to understand what traits have changed throughout their history, how often and in what order. The tree was constructed using a 600bp fragment of 28s rDNA. This tree suggests that the Epyrinae subfamily is in fact two distinct monophyletic clades, and should be split. Furthermore the Epyrini tribe gave rise to the Mesitinae and should be merged as a single taxon, cladistically speaking. The tree suggests a single evolution of tolerance at the base of the bethylids, a single major switch in host orders (Bethylinae attack Lepidoptera, the rest Coleoptera), convergent morphology between the Mesitinae and Chrysididae, and is consistent with the biogeography of the Mesitinae, which is the youngest major clade and is also the only group not found in both old and new worlds.
 see:
see:
Mayhew P.J. et al. in prep. Phylogeny and evolution of bethylid wasps inferred from D2 and D3 28s rDNA.
Top of the page
The Insect Parasitoid Special Interest Group of the Royal Entomological Society
The IPSIG aims to promote and disseminate research on insect parasitoids. The group is especially active in the study of behaviour, development, ecology, evolution and taxonomy, although we are keen to promote other fields. The group works informally to promote interaction and contacts via an e-mail list, updated annually. It also meets formally for a day-long annual workshop, usually in the spring, where new research is presented through talks and posters. The meeting often includes a longer presentation by an invited speaker and a small prize competition for the best oral presentation by a student. The meeting is attended by 40-50 people, mostly from the UK and neighbouring European countries.
Please contact me if you are interested in joining the e-mailing list.
The programme for the 2001 meeting, held in March, is below.
Top of the page
RES Insect Parasitoid Special Interest Group
Monday 26th March 2001, 41 Queens Gate, London SW7
Registration and coffee.
9.45-10.25. Registration is £2.00 for RES members and £5.00 for non-members. Non members can join up on the door if they wish. If you can arrive before 10.30, that will ensure a prompt start, since there is usually a queue to register!
10.25-10.30
Pete Mayhew (Chair) (York)
Introduction and welcome.
10.30-10.50
Becky Morris (Silwood Park)
"Spatial patterns of herbivore and parasitoid attack in a dry tropical forest: implications for the maintenance of tropical diversity"
10.50-11.10
Emilio Guerrieri (Portici, Italy)
"Plant-to-plant interactions: a successful strategy to attract insect parasitoids"
Coffee 11.10-11.40
11.40 - 12.00
John Pexton (York)
"The consequences of parent-offspring conflict upon life-history traits in parasitoids"
12.00 - 12.20
Shahab Manzari (Silwood Park)
"Determining species boundaries among whitefly parasitoids using morphometrics and DNA sequencing"
12.20 - 12.40
Daniel Bean (Silwood Park)
"Male mating tactics and lethal combat in a parasitic fig wasp"
Lunch
12.40-14.30. Lunch is not provided, but there are numerous pubs, restaurants and sandwich bars locally.....just follow the crowd!
14.30- 15.15
Invited speaker
Dr James Cook (Silwood Park).
"Fig wasp evolution and speciation - views from up above and down under"
15.15-15.35
Robin Williams (Wedmore, Somerset)
"British Oak Galls and their inhabitants."
TEA: 15.35-16.05
16.05-16.10.
Presentation of student prize
16.10-16.30
Richard Bailey (Edinburgh)
"Quantitative food webs and phylogenies"
16.30 - 16.50
Peter Mayhew (York)
"Molecular evidence for the phylogeny of the Bethylidae"
16.50-17.10
Graham Hopkins (Bristol)
"Leafminers and parasitoids in a tropical dry forest: foodweb structure"
17.10
Pete Mayhew
Closing remarks and notices, followed by drinks and posters
Drinks and Posters:
Vicky Schneider (Leiden)
"Geographic distribution and genetic relatedness in sexual and parthenogenetic
Venturia
canescens"
Bart Pannebakker (Leiden)
"Mutation accumulation and ecological genetics of sex in a haplodiploid
system"
Gijs Hoogendoorn (Leiden)
"Life-history and foraging behaviour evolution in Asobara persimilis
(Hymenoptera: Braconidae): selection along a geographic cline."
Top of the page
